Distribuição das imunoglobinas
No soro humano
IgM 10% IgGs (em conjunto) 70 a 75% IgA1 15 a 20%
Nos tecidos
IgM sangue (não atravessa o endotélio senão em
locais inflamatórios; não atravessa a placenta)
mIgM linfócitos B (membrana)
IgGs sangue (atravessam a placenta) e outros tecidos conjuntivos
IgA1 sangue
IgA2 secreções (inclusivamente glândulas mamárias),
sob a forma de dímero (IgA das secreções ou sIgA)
IgD linfócitos B (membrana)
IgE locais inflamatórios
Por funções
Efectoras
| Classe/ |
Complemento |
Fagocitose |
ADCC |
|
| Subclasse |
via clássica |
via FcR |
via FcRa |
Neutralização |
| IgM |
SIM |
|
|
SIM |
| IgG1 |
SIM |
FcgRI |
(FcgRIIa / FcgRIII) |
SIM |
| IgG2 |
|
|
|
SIM |
| IgG3 |
SIM |
FcgRI / FcgRIIa |
FcgRIIa / FcgRIII |
SIM |
| IgG4 |
|
FcgRI |
|
SIM |
| IgA1 |
|
FcaR |
|
SIM |
| sIgA |
|
|
|
SIM |
| IgE |
|
|
FceRI |
SIM |
a ADCC: citotoxicidade dependente de anticorpos; FcgRs
presentes em linfócitos K (IgG1 tem menos afinidade que IgG3); FceRI
em eosinófilos
Comentário: a função de neutralização
das IgG2 e das sIgAs, por ser a única que se conhece nelas, pode
ter a importância de não provocarem efeitos secundários
como por exemplo reacções de hipersensibilidade; a localização
das duas evidencia uma possível complementaridade entre elas. Mas
também pode significar, trivialmente, que outras funções
destas duas classes são apenas desconhecidas.
Reguladoras (activação das células do sistema
imunitário)
Formas de membrana nos linfócitos B
mIgM IgD
Via FcRs
IgM::Fc R linfócitos B
IgG3::Fc RII plaquetas
IgE::Fc RI mastócitos e basófilos, plaquetas, APCs excepto
B (receptor é induzido por citoquinas)
IgE::Fc RIIa linfócitos B
IgE::Fc RIIb APCs excepto B (receptor é induzido por citoquinas)
Alguns termos e conceitos em Imunologia
Resposta imunitária adquirida (ou adaptativa) vs. inata
-- Os mecanismos de reconhecimento específico de um determinado
antigénio (via imunoglobulinas e TcRs), porque conferem ao organismo
uma maior eficiência de protecção após o primeiro
contacto, definem a resposta adquirida ou adaptativa. Os mecanismos
que, envolvendo ou não os linfócitos B e T, não passam
por um reconhecimento específico e estabelecimento de maior eficiência,
são de natureza "geral" e definem a resposta inata. Esta
distinção é inconveniente por dois motivos: em primeiro
lugar, porque mesmo os mecanismos ditos inatos se encontram "adaptados"
a reconhecerem entidades químicas nonself, como por exemplo
as proteínas inflamatórias que ligam o manano (as ManBPs),
que suplementam as imunoglobulinas na activação do complemento
pela via clássica; o efeito adjuvante de certos produtos bacterianos
que lhes são específicos, como são os casos do lípido
A ou dos componentes da parede das micobactérias também revela
uma adaptação análoga embora se refira a resposta
adaptativa como uma adaptação do indivíduo e não
da população, é um termo que provoca confusão.
Em segundo lugar, porque deixa implícita uma separação
da resposta imunitária em vias "alternativas", quando na realidade
aquilo que se entende por adquirido envolve mecanismos inatos: nas resposta
primárias, o envolvimento dos mecanismos de reconhecimento específico
produtores de memória é desencadeado pela persistência
do antigénio após as fases mais imediatas ("inatas") da resposta.
Imunidade humoral vs. imunidade mediada por células
-- Refere, respectivamente, uma componente efectora da resposta imunitária
que é dependente de anticorpos e outra componente ligada a mecanismos
de citotoxicidade. Mas trata-se de uma extrapolação a partir
da definição "operacional" feita originalmente. A imunidade
humoral é aquela que pode ser transferida com o soro de indivíduos
imunes para indivíduos não-imunes: o organismo onde se injecta
o soro imune (o soro é um humor) "adopta" a competência
imunitária do dador e dado que essa competência é específica
considera-se que é mediada por anticporpos. A imunidade mediada
por células, em experiências de transferência adoptiva,
atribui-se em regra a linfócitos Tc presentes na população
celular transferida. Mas nas situações "reais", e dada a
interrelação funcional entre diversas classes de imunoglobulinas,
derivados do complemento, redes de citoquinas e de APCs, etc., nas respostas
imunitárias, é de pouca valia estabelecer uma distinção
entre o que depende de anticorpos e o que é mediado por células.
Afinidade vs. avidez -- As interacções entre
antigénios e anticorpos são comparáveis às
interacções enzima-substrato, definindo-se para cada tipo
de anticorpo e o antigénio que ele reconhece uma afinidade,
medida em unidades de l/mol (M-1). As IgMs têm em geral
uma bai«xa afinidade (Ka da ordem dos 107 M-1),
enquanto as IgGs têm geralmente uma alta afinidade (109
M-1). Mas a medição das constantes de afinidade
Ka requer antigénio e anticorpo purificados, o que no
caso destes últimos só é possível com anticorpos
monoclonais. Para soros policlonais, em vez de se falar de afinidade fala-se
de avidez, que é uma medida global da força das interacções
entre o antigénio (purificado) e os diversos anticorpos que o reconhecem,
presentes nesse soro (em condições experimentais padronizadas).
Se for viável extrapolar dos resultados experimentais para situações
in vivo, a avidez tem um grande significado fisiológico, pois permite
avaliar a imunidade do indivíduo para o antigénio com que
se fez a medição. Nestas situações, a avidez
mede-se, ao contrário da afinidade que é em medidas universais,
em termos relativos, sendo o padrão o soro pré-imune: do
mesmo indivíduo ou estirpe quando sem contacto prévio com
o antigénio. A avidez depende não só da afinidade
das CDRs de uma imunoglobulina mas também do número de CDRs
por imunoglobulina: as IgMs (pentaméricas) têm uma avidez
da mesma ordem de grandeza que as IgGs (monoméricas), formando redes
de interacções antigénio-anticorpo que atingem a zona
de equivalência com títulos semelhantes, apesar da sua mais
baixa afinidade, às IgGs.
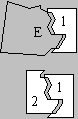 Idiótipo
vs. paratopo -- O idiótipo de um anticorpo é a
porção deste característica do clone de linfócitos
B que o produziu, e é formado pelo primeiro domínio conformacional
(a contar da extremidade amino) de um par de cadeias H e L em conjunto.
É ao nível do idiótipo que se define a especificidade
do anticorpo por um determinado antigénio, pois medeia a interacção
daquele com uma determinada conformação à superfície
do mesmo, o epitopo. O paratopo é a superfície de
contacto do idiótipo com o epitopo, a que lhe é tridimensionalmente
complementar; através das ligações não-covalentes
que se estabelecem entre estas duas superfícies que dá-se,
a nível molecular, o reconhecimento do antigénio. No caso
particular de um anticorpo anti-idiotípico (anticorpo 2 na figura),
que reconhece como epitopo o idiótipo de um outro anticorpo (1),
se for exactamente complementar do paratopo deste último verifica-se
que o paratopo 2 é uma réplica muito fiel do epitopo (E)
que é reconhecido pelo anticorpo 1.
Idiótipo
vs. paratopo -- O idiótipo de um anticorpo é a
porção deste característica do clone de linfócitos
B que o produziu, e é formado pelo primeiro domínio conformacional
(a contar da extremidade amino) de um par de cadeias H e L em conjunto.
É ao nível do idiótipo que se define a especificidade
do anticorpo por um determinado antigénio, pois medeia a interacção
daquele com uma determinada conformação à superfície
do mesmo, o epitopo. O paratopo é a superfície de
contacto do idiótipo com o epitopo, a que lhe é tridimensionalmente
complementar; através das ligações não-covalentes
que se estabelecem entre estas duas superfícies que dá-se,
a nível molecular, o reconhecimento do antigénio. No caso
particular de um anticorpo anti-idiotípico (anticorpo 2 na figura),
que reconhece como epitopo o idiótipo de um outro anticorpo (1),
se for exactamente complementar do paratopo deste último verifica-se
que o paratopo 2 é uma réplica muito fiel do epitopo (E)
que é reconhecido pelo anticorpo 1.
CD (cluster of differentiation) -- Conjunto de anticorpos monoclonais
(de murganho quase sempre) que reconhecem epitopos de uma molécula
cuja presença à superfície de certas células
do sistema imunitário contribui para a discriminação
(a sua diferenciação) em relação a outros
tipos celulares. É assim que, por exemplo, se podem distinguir diferentes
graus de activação na linhagem macrofágica, diferentes
subgrupos ("subsets") de linfócitos, etc.. O termo CD3 engloba muitos
anticorpos monoclonais específicos para um complexo proteico multimérico
associado ao TcR dos linfócitos T. Uma célula CD3+
é definida pela detecção desse complexo através
de um anticorpo CD3 e, por uma assimilação de linguagem,
também se designa a molécula reconhecida por CD3. O receptor
para a transferrina (marcador de linfócitos T activados, por exemplo)
é reconhecido pelos anticorpos CD71, pelo que por vezes é
também designado CD71.
Restrição MHC -- O reconhecimento dos antigénios
pelos linfócitos T é feito sobre pequenos péptidos
associados a moléculas MHC, o que implica terem os respectivos TcR
primordialmente uma especificidade para essas moléculas MHC. Diz-se
por isso que o antigénio activa um determinado clone T "no contexto
de" esse MHC. Verifica-se que, provavelmente pela selecção
negativa ao nível do timo, apenas entram na circulação
linfócitos T capazes de reconhecer os tipos MHC do próprio
(self), dado que as moléculas que vão apresentar os antigénios
são do mesmo indivíduo; por isso a restrição
MHC é primordialmente ao MHC self. No entanto, na rejeição
de enxertos, na doença da reacção do enxerto contra
o hospedeiro (GVHD), ou no modelo experimental das culturas mistas de linfócitos
(as MLC, mixed lymphocyte cultures), o reconhecimento (e reacção
citotóxica subsequente) é feito num contexto MHC nonself
(alo-reactividade), interpretável como resultante de uma
reactividade cruzada entre as especificidades existentes (restritas ao
MHC self) e certas especificidades em contextos nonself.
[Extraído de
um manual de toxicologia, onde entre outras coisas se afirma que apenas
o valor da LD50 é insuficiente para informar sonre
o poder toxígeno de uma substância, para tal sendo necessário
pelo menos relatar outros dados experimentais]
Condições que afectam as condições de toxicidade
de qualquer composto
Dose -- em geral, quanto maior é mais rápida
é a observação dos efeitos.
Via de administração -- em geral, a toxicidade
é máxima pela via que leva mais rapidamente a substância
tóxica até à circulação sanguínea:
i.v. (intra-venoso) > inalação > i.p. (intra-peritonial)
> i.m. (intra-muscular) > s.c. (sub-cutânea) > oral > cutânea
(intra-dérmica ou i.d.)
Regime e local de injecção -- a quantidade
de material tóxico injectado por minuto influencia consideravelmente
a dose tóxica efectiva; também a escolha da veia por onde
se injecta pode (dependendo dos obstáculos e tempo que se interpõem
entre o local de injecção e os tecidos-alvo) influenciar
a dose tóxica efectiva.
Uma medida alternativa à LD50 é a obtenção
de intervalos de confiança estatística de toxicidade, geralmente
de 95%, a partir do ajustamento de uma curva matematicamente conhecida
(por exemplo a curva de Gauss cumulativa). Estes intervalos são
muito mais informativos, mas requer que os dados tenham sido obtidos a
uma escala suficientemente grande.
Uma classificação dos compostos por toxicidade coloca
o nível de "extrema toxicidade" (classe 1), para a administração
via oral em murganhos, a < 1 mg / Kg de peso.