Embora o cariótipo seja em princípio um carácter unificador de cada espécie de ser vivo, o reconhecimento de variações a nível cromossómico pode ter grande interesse. Na constituição cromossómica de cada indivíduo está a descrição citológica do seu genótipo, sendo pois as variações na constituição cromossómica uma fonte importante de informação explicativa de variações (frequentemente, anomalias) a nível fenotípico, isto é: certos fenótipos "mutantes", por serem consistentes com certos padrões de variação cariotípica, serão provavelmente causados por essas variações.
Ao longo do ciclo de vida, reconhecem-se fases haplóides (n), diplóides (2n) e triplóides (3n), características respectivamente dos gametófitos, esporófitos e endosperma; certos tecidos do esporófito podem apresentar múltiplos dos 2n cromossomas (poliploidia) em resultado da endoduplicação, processo amplificador da informação genética na célula em que entre sucessivas fases S não há mitoses. Mas o cariótipo representativo da espécie é sempre (em plantas superiores) o diplóide.
EspeciaisCertos organismos são fruto da endoduplicação após uma hibridação interspecífica: são alopoliplóides. Tomando como exemplo o trigo mole (Triticum æstivum), a sua fórmula cariotípica é dupla: os 42 cromossomas tanto correspondem à fórmula diplóide 2n = 42 cromossomas, como à fórmula hexaplóide 6x = 42, servindo esta última para especificar que existem três conjuntos diplóides. A diferença está no número básico de cromossomas, que não é o haplóide (que segrega nos gâmetas) n = 21, mas sim x = 7 cromossomas, agrupados por três cariótipos que se designam por letras (A, B, D) e provêm de T. durum (A e B), outra espécie alopoliplóide (2n = 4x = 28) e T. tauschii (D), diplóide; os progenitores do hexaplóide terão sido um alotetraplóide AABB e um diplóide DD que (segundo parece há 10000 anos) se hibridaram, resultando num zigoto triplóide ABD que por sua vez se desenvolveu no hexaplóide AABBDD por endoduplicação. Os sete pares A, B e D são sinténicos entre si, isto é, contêm genes semelhantes e no mesmo ordenamento (cf. "mapeamento"), o que é de esperar entre espécies cujo conteúdo genético deve ser muito semelhante; mas durante a meiose os cromossomas A B e D portam-se entre si como cromossomas não-homólogos: os cromossomas A emparelham e segregam só com cromossomas A, idem B com B e D com D, mas não A com B, etc.. Em resultado disso, cada gâmeta mantém um conjunto completo de 7 cromossomas de cada, A, B e D. Devido a estas características, passaram a designar-se homeólogos os cromossomas que contêm informação genética semelhante mas não-homóloga: os dois cromossomas 1A de um determinado indivíduo são homólogos um do outro, mas homeólogos dos 1B e dos 1D, e assim sucessivamente.
A formação de alopoliplóides não é de todo infrequente na natureza, e pode ser induzida experimentalmente como sucedeu no caso do Triticale (trigo × centeio). E conhecem-se casos férteis fora dos cereais, como na cana-do-açúcar, no nabo e no tabaco (alopoliplóides naturais) e soja (alopoliplóides sintéticos).
Os autopoliplóides podem resultar de endoduplicação a partir de um zigoto diplóide, ou (o que é particularmente frequente em Solanum spp.) da não-segregação das cromátides na segunda divisão da meiose, produzindo gametófitos diplóides cujos gâmetas se conjugam para formar autotetraplóides.
Na meiose, tanto na anafase I como na II podem surgir erros na distribução dos cromossomas, de tal maneira que ocasionalmente o que devia segregar migra conjuntamente para o mesmo pólo da célula: trata-se de não-disjunções, e o seu resultado são gâmetas com n + 1 cromossomas (e, reciprocamente, n – 1, caso o gametófito sobreviva à ausência de um dos cromossomas); este é um dos mecanismos que se sabe poderem estar na origem de esporófitos aneuplóides: ao contrário das constituições cromossómicas que têm múltiplos inteiros do número básico n da espécie (euploidias), nas aneuploidias há um desequilíbrio entre as doses genéticas de diferentes grupos de ligação. Tal desequilíbrio traduz-se num fenótipo facilmente reconhecível morfologicamente, sendo utilizado como complemento da análise mendeliana para o estabelecimento de uma correspondência entre loci de interesse e o respectivo grupo de ligação.
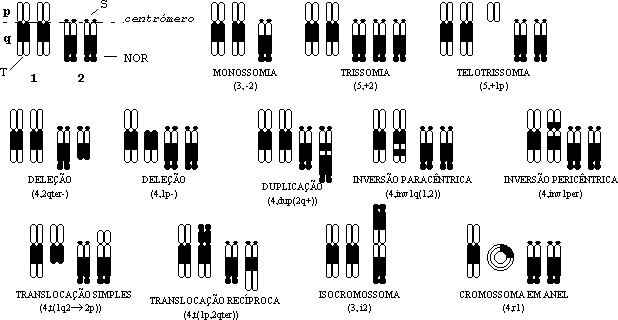
Diz-se que há uma monossomia quando falta um cromossoma dum par de homólogos, e uma trissomia ou tetrassomia se tiver três ou quatro (figura 5); o isolamento de nulissómicos, especialmente em alopoliplóides, tem permitido analisar a contribuição específica de um par de cromossomas, para os diversos fenótipos em estudo, em relação aos seus homeólogos.
Os cromossomas podem sofrer alterações estruturais de diversa ordem, com diversas implicações nos fenótipos dos indivíduos ou na sua fertilidade. A figura 5 servirá para ilustrar esquematicamente algumas das variações, numéricas e estruturais, que mais usualmente se encontram:
Figura 5 — Constituições cromossómicas hipotéticas (n = 2) em metafase, ilustrando o cariótipo normal e exemplos de variações numéricas e estruturais, evidenciadas por bandeamento, e nomenclatura utilizada para as designar. No cariótipo normal, com dois grupos de ligação (1 e 2), legendam-se as diversas partes dos cromossomas; NOR: região organizadora do nucléolo; T: telómero; S: satélite; p, q: braços dos cromossomas. Cada fórmula cromossómica contém primeiro o número total de cromossomas, seguido de uma lista das anomalias, com os cromossomas identificados pelos respectivos números.
As consequências fenotípicas das anomalias estruturais derivam de um efeito de dose atribuível (como nas aneuploidias) ao número incorrecto de certos genes (a mais nas duplicações e a menos nas deleções), ou de um efeito de posição resultante da presença de um locus numa localização anormal (inversões, translocações, cromossomas em anel).
As translocações Robertsonianas têm a particularidade de envolverem a fusão ou cisão de cromossomas ao nível do centrómero, por exemplo a criação de um cromossoma metacêntrico a partir de dois acrocêntricos. É ocorrência relativamente comum na natureza espécies aparentadas terem cariótipos muito diferentes por via de translocações deste tipo, sendo muito plausível que a separação inicial entre as espécies tenha em tais casos envolvido rearranjos que isolem reprodutivamente dois grupos, como se explica a seguir.
Problemas de fertilidadeOs cromossomas na profase I da meiose, mais precisamente a partir do zigóteno e até ao paquíteno, passam por um processo de "busca de homologias" mútuas, culminando no correcto emparelhamento que assegura o crossover absolutamente recíproco entre os homólogos. Se houver discrepâncias estruturais entre os homólogos, podem resultar bivalentes anómalos, ou tetravalentes, como se ilustra na figura 6:
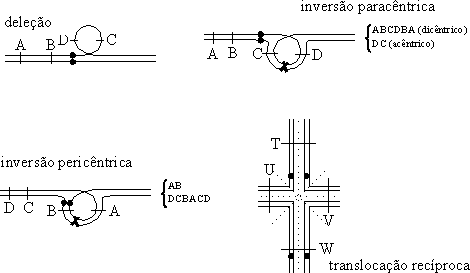
Figura 6 — Bivalentes e tetravalentes formados entre cromossomas homólogos com discrepâncias estruturais. Nas inversões exemplificam-se crossovers e (nas chavetas) os cromossomas resultantes após a meiose (cf. texto).
Como indicado, a ocorrência de crossovers nas "ansas" das inversões vai produzir cromossomas anormais, acarretando gametófitos desequilibrados geneticamente, muitas vezes abortivos:
i) um crossover dentro da inversão pericêntrica vai resultar em dois cromossomas que segregam deleções e duplicações;
ii) um crossover dentro da inversão paracêntrica vai resultar num cromossoma acêntrico e num cromossoma dicêntrico, ambos com deleções do material genético, que se encontra duplicado no homólogo; o cromossoma acêntrico perder-se-á no decurso das divisões celulares, e o dicêntrico tenderá a migrar simultaneamente para os dois pólos na anafase I, partindo-se em dois durante a telofase seguinte, sempre em pontos imprevisíveis, de tal modo que as células filhas são sempre únicas geneticamente, deficientes (na fase haplóide) e desequilibradas; adicionalmente, os cromossomas que se partiram não têm telómeros, e assim as extremidades podem "aderir" ao outro braço do mesmo cromossoma e produzirem um cromossoma em anel;
iii) um número ímpar de crossovers envolvendo um cromossoma em anel vai resultar num único cromossoma dicêntrico, de que poderá resultar um desequilíbrio genético (e novamente um cromossoma em anel) como descrito no parágrafo anterior.
No caso das translocações, robertsonianas ou recíprocas, cria-se um problema na resolução dos tetravalentes na anafase I, em que de modo análogo à resolução dos quiasmas é necessário que se efectuem quebras que permitam a migração dos cromossomas para os pólos da célula. Na figura 6, ilustram-se a tracejado as hipóteses de resolução numa translocação recíproca, uma das quais até repondo a ordem normal.
|
Também há problemas de fertilidade em muitos alopoliplóides, ou porque há ambiguidade de emparelhamentos (tetravalentes) ou porque as meioses são abortivas. |