Na sua qualidade de unidades estruturais de transmissão do material genético, os cromossomas têm uma dupla faceta, de estabilidade e dinamismo, ambos necessários à correcta expressão dos genes: estabilidade no conteúdo dessa informação e seu arranjo, com uma baixíssima taxa de mutação nos processos replicativos (apesar das inúmeras cópias que a partir do zigoto se produzem para o desenvolvimento de um organismo multicelular, a taxa de mutação por geração permanece sempre a níveis que não alteram significativamente a 1ª lei de Mendel); e dinamismo, representado por três processos integrantes das divisões celulares, designadamente a duplicação da informação genética, visível nas profases e metafases pelas duas cromátides unidas ao nível do centrómero de cada cromossoma, a condensação das cromátides durante as profases, pela qual os diferentes cromossomas se "desemaranham" do entranhado em que se encontravam no núcleo interfásico, e, especificamente na meiose, o crossover.
A interfase abrange todos os processos desde o final de uma mitose ao eventual início da mitose seguinte; o núcleo e os nucléolos encontram-se bem individualizados, enquanto os cromossomas estão descondensados ao ponto de ser muito complexa a sua distribuição dentro do núcleo, não podendo ser observados directamente senão em condições ou em casos muito especiais.
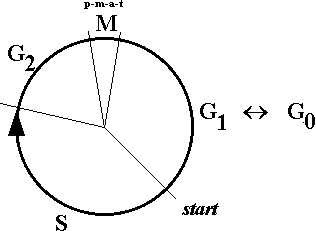
A célula, à saída da mitose, entra em interfase e pode nunca mais voltar a dividir-se, mas se voltar inicia o processo preparando-se para a replicação do DNA, que dura várias horas e produz no final, a partir de cada molécula desse DNA, duas moléculas idênticas entre si (as duas cromátides) e idênticas à molécula que lhes antecedeu (cf. "replicação"). Como mostra a figura 3, este período da interfase é denominado a fase S (síntese do DNA), e entre ela e o início da profase seguinte há um intervalo de duração mais ou menos característica em cada tipo de célula, a fase G2 (G de gap, sinónimo inglês de intervalo, lacuna).
Figura 3 — Diagrama das diferentes fases do ciclo celular: M (mitose, subdividida nas sucessivas fases pelas correspondentes iniciais), G1 (gap1, pré-síntese do DNA), S (replicação do DNA), G2 (gap2, pré-mitose); a sequência G1——S——G2 constitui a interfase. O ponto "start" indica a transição G1 → S que marca a "decisão" da célula de se dividir. G0 não é uma fase, é um estado de não-divisão celular, reversível ou não para G1 (eventualmente G2).
A entrada na profase marca o início da condensação dos cromossomas, que vai atingir o seu máximo na metafase e anafase. Dado que a metafase é relativamente prolongada e os cromossomas são mais fáceis de individualizar entre si, as imagens mais comuns referem-se a esta fase da mitose; deste modo, os cariótipos — o tipo citológico do material nuclear — das espécies eucariotas definem-se pelo menos em função da observação de cromossomas metafásicos (cf. figura 5): no máximo da sua condensação e contendo duas cromátides.
A região onde aparentemente se unem as cromátides de cada cromossoma metafásico é a constrição primária ou centrómero, e é também por essa região que as cromátides vão ser puxadas para cada um dos pólos da célula durante a anafase (por isso os pedaços de cromossoma sem centrómero, ou cromossomas acêntricos, estão condenados a perder-se na sequência das divisões celulares). É por referência ao centrómero que se definem os caracteres morfológicos principais de cada cromossoma: se a sua posição for terminal ou quase, o cromossoma diz-se acrocêntrico (ou telocêntrico) e tem apenas um braço por cromátide. Sendo medial, o cromossoma é metacêntrico e tem dois braços por cromátide, prolongando-se em sentidos opostos, distinguindo-se um braço mais curto (p) do mais longo (q).
Os telómeros são os segmentos terminais de cada braço, e desempenham um papel importantíssimo na estabilidade dos cromossomas, durante o ciclo celular nomeadamente.
Em cada cariótipo há pelo menos um par de cromossomas com outra constrição ("secundária") que contém a região organizadora do nucléolo (NOR), dado que é à sua volta que o nucléolo se reconstitui na telofase. Estruturas conspícuas em muitos cariótipos são os satélites, material por vezes de dimensão variável que se prolonga a partir do centrómero de cromossomas acrocêntricos, ou constitui uma região bem definida no extremo de um braço (contendo, assim, o respectivo telómero).
Quando se detecta uma célula em metafase, num esfregaço devidamente preparado e corado, a imagem que se apresenta é a de cromossomas espalhados que se fotografam, recortando-se as figuras individualizadas dos cromossomas para serem agrupadas duas a duas (por pares de homólogos, em princípio de morfologia idêntica), e em seguida serem ordenadas e numeradas por tamanho, desde os maiores aos mais pequenos.
Com bastante prática é possível distinguir, nas espécies que se estuda, os
diferentes pares de cromossomas, mas existem casos onde é necessário um
tratamento (nas plantas, geralmente, o bandeamento C) que resulta na
coloração diferencial da cromatina em diferentes segmentos de cada braço,
evidenciando padrões de bandeamento característicos de cada par
cromossómico; assim se consegue a identificação de cada par sem
ambiguidades. O bandeamento C também é utilizado para distinguir cromossomas
homeólogos (v. adiante).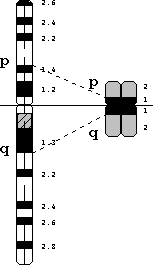
Nas plantas não se têm explorado pormenores de bandeamento cromossómico ao ponto a que tem sido feito na espécie humana e nalguns animais; pode ir-se ao ponto de analisar os cromossomas pré-metafásicos, que por estarem ainda em processo de condensação são mais compridos mas já suficientemente espessos para a observação microscópica; a estrutura de bandas observáveis na metafase decompõe-se em sub-bandas nestes cromossomas, podendo ser de grande auxílio na identificação de variações estruturais (figura 4).
Figura 4 — Comparação entre o cromossoma 3 humano em pré-metafase (esquerda) e em metafase; a maior distensão na pré-metafase permite distinguir sub-bandas dentro das bandas principais visíveis em metafase (neste caso, com bandeamento G). A linha horizontal indica a posição do centrómero nestas imagens.
Na profase I há dois aspectos citológicos de interesse, e que estão especialmente bem estudados no milho: os cromómeros e os pontos de quiasma. Os cromómeros são observáveis desde o leptóteno ao paquíteno, constituindo zonas mais condensadas dos cromossomas intercaladas por "espaçadores" menos condensados, dando-lhes um aspecto de "colares de pérolas". Estes cromossomas fornecem a informação estrutural mais detalhada em plantas, quase ao nível de resolução dos cromossomas politénicos dos dípteros.
Os pontos de quiasma observam-se particularmente bem a partir do diplóteno, permanecendo até à anafase I, altura em que se dá a sua "resolução" que liberta os cromossomas homólogos em início de segregação. Há evidência muito segura de haver uma correspondência directa entre o número de pontos de quiasma e o número de crossovers que permitem a recombinação entre cromossomas homólogos (cf. "grupos de ligação"), pelo que estas figuras citogenéticas são consideradas evidência directa dessa recombinação.