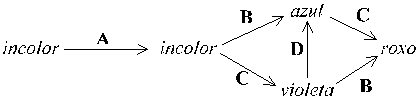
Todo o fenótipo resulta da acção de uma série de genes envolvidos no seu desenvolvimento; se dizemos que um determinado indivíduo é mono-híbrido ou di-híbrido, é apenas porque a variação genotípica que ele contém, correspondente às diferenças entre os seus progenitores, envolve um ou dois loci, respectivamente. Por isso há muitas experiências em que a variação num único fenótipo em estudo resulta de mais de um par de genes alelos, podendo neste caso detectarem-se interacções entre os genes não-alélicos, portanto entre diferentes loci; nesses casos, a relação um locus ↔ um fenótipo (por exemplo um locus determinando a cor dos cotilédones e outro a forma da semente, como Mendel verificou) não se aplica, e ainda assim as duas leis de Mendel são verdadeiras: as ditas leis referem-se unicamente à transmissão dos genes pelos gâmetas, não às relações que estes estabelecem entre si para estabelecerem os fenótipos.
|
Em certos caracteres, ditos poligénicos (cf. "variação contínua"), o hibridismo envolve tantos loci que a individualização de classes fenotípicas atribuíveis a determinado(s) genótipo(s), na segregação a partir dos híbridos, se torna impossível, obrigando à análise estatística das populações segregantes. |
As situações em que dois ou mais loci determinam em conjunto um único fenótipo só merecem designação especial quando os genes de loci diferentes tenham modos de interacção que implicam o agrupamento de genótipos, donde uma distribuição por classes na F2 (tanto a partir do selfing como ainda a partir do testcross) diferente do que já foi caracterizado na secção anterior. Descobriram-se logo a partir da 1ª década do século XX, em fenótipos cuja variação era determinada por mais de um locus, vários tipos de interacções entre loci, a que muitos autores apelidam colectivamente de epistasia, se bem que seja uma designação que só se aplica em rigor a alguns. Seguem-se exemplos no caso mais simples, que é o de dois loci de segregação independente, com dominância completa dos genes A e B.
Se os genes A e B tiverem o mesmo efeito fenotípico, sendo indiferente estar presente um ou o outro, ou até os dois, dizem-se redundantes ou duplicados, e a segregação na F2 é de 15 : 1 (selfing) ou 3 : 1 (testcross). Nilsson-Ehle (Escandinávia, 1909) apresentou um exemplo destes no cruzamento entre trigo de grão vermelho e grão branco: na F2 (F1 × F1, em que a F1 tinha grão vermelho), apenas 1/64 dos grãos eram brancos, correspondendo provavelmente a um genótipo do tipo aabbcc (redundância entre os genes dominantes A, B e C).
Se os genes A e B só produzem um efeito fenotípico quando presentes em conjunto, dizem-se complementares ou sinergísticos, obtendo-se uma segregação na F2 de 9 : 7 (selfing) ou 1 : 3 (testcross).
Uma situação duplamente de redundância e de complementaridade corresponde a quando os genes A e B, isolados, produzem um efeito fenotípico igual, formando-se uma classe reunindo os genótipos das classes A-bb e aaB-, mas diferente da classe A-B-, onde eles se complementam (9 : 6 : 1 no selfing, 1 : 2 : 1 no testcross).
A epistasia define, ao contrário das anteriores, uma hierarquia entre os dois loci: quando um certo genótipo de um locus estabelece por si só o fenótipo, independentemente do genótipo no outro locus, diz-se que o primeiro é epistático sobre o segundo, sendo o que fica "mascarado" hipostático. Considerando ainda o caso mais simples, existem duas possibilidades:
i)o efeito epistático resulta da presença de um gene dominante (digamos que é o A): resultam três classes, A---, aaB- e aabb, em que a variação no locus B/ b só é aparente na ausência do gene epistático A (12 : 3 : 1 no selfing, 2 : 1 : 1 no testcross; variante: quando o fenótipo de A--- é igual ao de aabb, produzindo duas classes fenotípicas apenas nas proporções de 13 : 3 e 3 : 1 respectivamente).
ii)o efeito epistático deriva de um gene recessivo (digamos que é o a): resultam três classes, A-B-, A-bb, e aa--, em que a variação no locus B/ b só é aparente na presença do gene dominante A no locus epistático (9 : 3 : 4 no selfing, 1 : 1 : 2 no testcross).
Exemplo
Em certos casos, através da dedução dos genótipos pela análise das populações segregantes, é possível estabelecer uma correspondência entre um determinado fenótipo e os passos do metabolismo necessários ao seu desenvolvimento. Por exemplo, considere-se a síntese dos pigmentos das pétalas resultante de uma determinada via metabólica cujos catalizadores enzimáticos são a expressão directa de genes que segregam para variações da cor das pétalas: a partir de um substrato incolor tínhamos o seguinte conjunto de reacções:
Figura 2 — Via metabólica hipotética de bio-síntese da pigmentação de uma flor. Cada reacção é catalizada por um de quatro enzimas (A–D), mas encontra-se bloqueada em plantas homozigóticas recessivas para o locus correspondente (por exemplo, todas as flores de genótipo aa são incolores). Seguem-se alguns exemplos de genótipos de plantas di-híbridas e os resultados que se esperariam do selfing em cada uma (com segregação independente dos 4 loci), e a sua interpretação:
Genótipo | Fenótipo | Segregação após selfing | Interpretação genética |
|---|---|---|---|
AABbCcdd | roxo | 9 roxo : 3 violeta : 3 azul : 1 incolor | sem interacção aparente |
AABbCCDd | roxo | 15 roxo : 1 violeta | B e D redundantes |
AabbCcDD | roxo | 9 roxo : 7 incolor | A e C complementares |
AABbCcDD | roxo | 12 roxo : 3 azul : 1 incolor | C epistático sobre B/b |
AabbCCDd | roxo | 9 roxo : 3 violeta : 4 incolor | a epistático sobre D/d |
Note-se como, neste exemplo, os dois híbridos BbCc exemplificados podem mostrar ou não um efeito epistático dependendo do genótipo num terceiro locus (D/d): as proporções obtidas entre as classes fenotípicas podem ou não denunciar interacções de acordo com os backgrounds genéticos prevalecentes (cf. "efeitos do ambiente").
Muitos autores utilizam o termo epistasia num sentido lato para expressar qualquer tipo de interacção não-alélica, se bem que ao fazerem-no para interacções como a de redundância e de complementaridade tenham de dizer que os dois loci são epistáticos um sobre o outro, com isso não respeitando o significado etimológico do termo, que implica hierarquia entre eles.